Древнее оледенение и жизнь
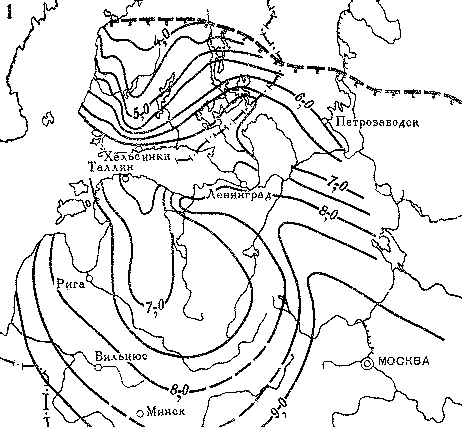
Для примера рассмотрим последовательность распространения ели обыкновенной — характерной древесной породы европейской тайги (рис. 19). Во время последнего оледенения она произрастала в более южных районах Русской равнины, а в центральных районах уступала место холодовыносливой сибирской ели, которая растет даже там, где существует многолетняя мерзлота.
Улучшение климата в самом конце ледниковой эпохи вызвало миграцию ели обыкновенной к северу, и около 10 — 8 тыс. лет назад она постепенно заселила центральные районы Русской равнины, хотя тогда она еще не играла видную роль в составе лесов. Одновременно ель сибирская отступила в северо-восточные районы. К концу указанного интервала времени ель обыкновенная проникла в восточную часть Прибалтики и на юг Онего-Ладожского перешейка.
За следующие полторы тысячи лет (8 — 6,5 тыс. лет назад) темпы расселения ели на севере уменьшились, и только на западе Прибалтики и на севере Польши распространение ели еще продолжалось. Именно в это время, по-видимому, произошло слияние двух миграционных потоков ели с юга, со стороны Карпат, и с востока, со стороны Русской равнины.
Во время послеледникового климатического оптимума (6,5 — 5,8 тыс. лет назад) появились наиболее благоприятные условия для увеличения ели в лесах и расширения ее ареала. Эта порода проникла в северные районы Эстонии, и тем самым завершился процесс ее распространения на северо-западной окраине Русской равнины.
На территории Финляндии и Швеции ель мигрировала очень быстрыми темпами. Примерно 3,6 — 3,3 тыс. лет назад эта порода появилась в Центральной Лапландии, а еще спустя несколько столетий достигла подножий Скандинавского нагорья. Расселение ели в центральные и южные районы Швеции произошло в самые последние тысячелетия и продолжается до сих пор.
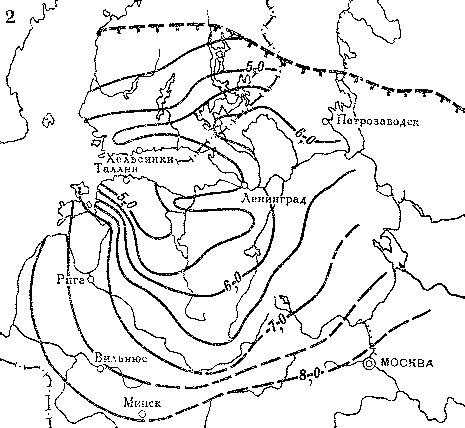
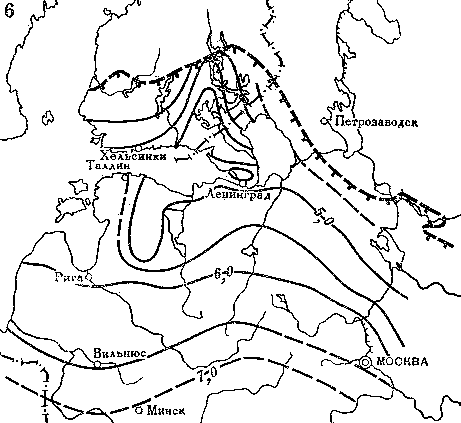
Приведенные факты показывают, что ель в послеледниковое время мигрировала из центральных районов Русской равнины на северо-запад. Для других пород в пределах той же территории были выявлены несколько иные пути расселения (рис. 20). Так, например, липа в послеледниковое время продвигалась двумя разными путями — на север вдоль восточного побережья Балтийского моря и на северо-запад из Подмосковья по возвышенностям в бассейнах рек Тверцы и Меты к побережью Финского залива. Спустя 2,5 тыс. лет оба миграционных потока соединились на севере Эстонии, тогда как на обширной промежуточной территории в бассейне Чудского и Псковского озер расселение липы происходило с большим запозданием.
Рис. 19. Распространение ели на востоке и севере Европы
1 — основной ареал ели около 8 тыс. лет назад; 2 — разреженные островные местообитания ели около 8 тыс. лет назад; 3 — граница современного ареала и островные местообитания ели европейской; 4 — граница современного ареала ели сибирской; 5 — изохроны появления ели (возраст по 14С указан в тыс. лет)
Несмотря на очень быстрые темпы распространения в послеледниковое время теплолюбивых широколиственных пород — липы, дуба, вяза, выяснилось, что даже во время климатического оптимума они не проникали далеко на север Европы, за пределы своих нынешних ареалов. Северных границ эти породы деревьев достигли около 4,5 — 4,0 тыс. лет назад, и с тех пор имели место только небольшие осцилляции. Такой вывод расходится с представлениями о продвижении теплолюбивых древесных пород на территории Сибири к берегам морей Северного Ледовитого океана во время послеледникового климатического оптимума и о последующем возвращении их в пределы современных ареалов.
Чтобы представить себе общую направленность миграций растительности на территории Русской равнины в голоцене, обратимся к исследованиям, выполненным в центрально-черноземных областях с применением спорово-пыльцевого и радиоуглеродного анализов.
Во время последнего плейстоценового оледенения на возвышенных участках этой территории распространялись перигляциальные травянистые сообщества в сочетании с зарослями ерника (кустарниковых берез) и ольховника. Структура растительного покрова отличалась мозаичностью и во многом зависела от пространственного распределения разных типов многолетнемерзлых грунтов. Однако по крупным долинам даже в это суровое время произрастали редкостойные таежные леса из сосны, ели, лиственницы и березы. Подобную ярусность растительности с лесами по долинным таликам и кустарниково-травянистыми сообществами на плакорах в настоящее время можно наблюдать на предгорных равнинах в Восточной Якутии.
Улучшение климата в конце плейстоцена вызвало таяние многолетней мерзлоты и распространение переувлажненных грунтов в перигляциальной зоне Русской равнины. В составе травянистых сообществ того времени было много осок. В более сухих местообитаниях в изобилии росли полыни и маревые. Значительно улучшились условия произрастания древесных пород, однако редколесья все еще концентрировались преимущественно по речным долинам. Основной фон растительного покрова составляли перигляциальные кустарниково-травянистые сообщества. Пространственная структура растительности претерпевала изменения под влиянием климатических пульсаций. Похолодания способствовали большему распространению открытых перигляциальных ландшафтов, потепления стимулировали расширение площадей редкостойных лесов.
Процесс перестройки перигляциальной растительности закончился в раннем голоцене, когда установилась широтная дифференциация растительных зон, близкая к современной. В бореальный период (9 — 8 тыс. лет назад) на северо-западе Среднерусской возвышенности произрастали сосновые и березовые леса. По направлению к югу и юго-востоку в лесах наряду с сосной и березой становилось больше широколиственных пород. На севере возвышенности среди этих пород в первую очередь появлялись липа и вяз, на юго-западе и западе главную роль играл дуб. Проникновение широколиственных пород происходило с запада и юго-запада, из Подольского рефугиума термофильной флоры. Южная граница лесной зоны в бореальное время пересекала центральную часть Среднерусской возвышенности по линии Железногорск — Кромы. К юго-востоку от этого рубежа возрастала роль лугово-степных сообществ, которые в сочетании с массивами лесов формировали зону лесостепи, сменявшуюся на юге возвышенности степной зоной.
Рис. 20. Распространение широколиственных пород на северо-западе СССР и юге Финляндии в послеледниковое время
Изохроны появления: 1 — липы; 2 — вяза; 3 — дуба. Изохроны начала широкого распространения в лесах: 4 — липы; 5 — вяза; 6 — дуба. Цифрами обозначены значения возраста (в тыс. лет) от 1950 г. Штриховыми линиями с подсечками выделены северные границы современных ареалов соответствующих пород
В атлантический период (7 — 5 тыс. лет назад) вся лесная зона на Русской равнине значительно продвигалась к северу, соответственно в том же направлении смещалась и степная зона. При этом лесостепная зона значительно сужалась, поскольку положение южной границы лесной зоны сравнительно мало менялось. Сходный вывод о стабильности южной границы лесной зоны в атлантический период сделан и для юга Западной Сибири.
Палеоботанические данные свидетельствуют, что в это время северные и западные районы Среднерусской возвышенности были заняты лесами с большой примесью широколиственных пород: на севере липы, на западе дуба. Вся южная половина возвышенности была занята степями, и северная граница степной зоны доходила до верховьев Оки.